
Allergy. 2017 Feb 22. doi: 10.1111/all.13145.
[Epub ahead of print]
Author information
- 1Institute of Pharmacology, University of Bern, Bern, Switzerland.
Abstract
 The network of cells and soluble mediators implicated in the pathogenesis of asthma and allergic disorders is complex. Deciphering details of the crosstalk between its components is essential for the identification of novel drug targets and for advances in patient management and precision medicine (1, 2). There is increasing evidence that innate lymphoid cells (ILCs) contribute to allergic responses (3). This article is protected by copyright. All rights reserved.
The network of cells and soluble mediators implicated in the pathogenesis of asthma and allergic disorders is complex. Deciphering details of the crosstalk between its components is essential for the identification of novel drug targets and for advances in patient management and precision medicine (1, 2). There is increasing evidence that innate lymphoid cells (ILCs) contribute to allergic responses (3). This article is protected by copyright. All rights reserved.
This article is protected by copyright. All rights reserved.
- PMID: 28226397
- DOI: 10.1111/all.13145
- [PubMed – as supplied by publisher]
-
Publication Types
Publication Types

PROF. STEPHAN VON GUNTEN (Orcid ID : 0000-0002-2753-8738)
Received Date : 15-Feb-2017
Accepted Date : 16-Feb-2017
Article type : Editorial
EDITORIAL
(on “CB2 receptors regulate natural killer cells that limit allergic airway inflammation in a murine model of asthma” for Allergy; ref number ALL-2016-00684.R1)Title:
Innate lymphoid cells in asthma: cannabinoids on the balance.
,
(Boligan KF, von Gunten S)
Institute of Pharmacology, University of Bern, Bern, Switzerland
Corresponding author: , MD, PhD, MME, Institute of Pharmacology,University of Bern, INO-F, Inselspital, CH-3013 Bern, Switzerland. Tel.: +41 31 632 32 98;Fax: +41 31 632 49 94; e-mail: stephan.vongunten@pki.unibe.chThis article has been accepted for publication and undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process, which may lead to differences between this version and the Version of Record. Please cite this article as doi: 10.1111/all.13145
This article is protected by copyright. All rights reserved.
Conflict of Interest: The authors declare no conflicts of interest.
Author contribution: All authors contributed with manuscript writing.
The network of cells and soluble mediators implicated in the pathogenesis of asthma and allergic disorders is complex. Deciphering details of the crosstalk between its components is essential for the identification of novel drug targets and for advances in patient management and precision medicine (1, 2). There is increasing evidence that innate lymphoid cells (ILCs) contribute to allergic responses (3). ILCs represent subsets of lymphocytes that express distinct patterns of cytokines and, similar to T and B cells, arise from a common lymphoid progenitor (CLP) cell (4). Given that natural killer (NK) cells, the prototypical member of the ILC family (4), express Fc-receptors, their activity depends on the composition of the individual antibody repertoire (5, 6), but is also tightly balanced by activating and inhibitory receptors (7). NK cell inhibitory receptors are specific for MHC class I, whereas others recognize non-MHC protein or glycan ligands (7, 8). Furthermore, Massi et al. reported that endogenous cannabinoids inhibit NK cell activity by engagement of the G protein-coupled cannabinoid receptors CB1 and CB2 (9). In contrast to NK cells, which have been classified as group 1 ILC, group 2 innate lymphoid cells (ILC2) are characterized by the production of T helper 2 (TH2) cell -associated cytokines (3), and increasing evidence demonstrates their contribution to allergic disorders, including asthma (reviewed in (4)). Recently, increased numbers of ILC2 cells in mucosal samples of patients with chronic rhinosinusitis (CRS) were shown to correlate with TH2 cell frequency (10), supporting the notion that interactions between ILC2 cells and the adaptive immune system might take place during allergic inflammation (4). Does crosstalk also exist between ILC subsets such as NK cells and ILC2 in allergic inflammation and by what factors would it be influenced?
In the current issue of Allergy, Ferrini et al. (11) provide evidence suggesting a role of NK cells in limiting ILC2 responses and subsequent allergic airway inflammation depending on NK cell regulation by CB2. Intranasal exposure to house dust mite (HDM) allergen led to reduced allergic airway inflammation in CB2-/-mice, a finding that was also observed when WT animals were treated with the CB2-specific antagonist AM630. Interestingly, CB2 -/- mice displayed elevated numbers of NK cells in the lungs, which was inversely correlated with ILC2 frequency. Depletion of NK cells in CB2 -/- mice restored airway inflammation and was associated with increased ILC2 numbers in the lung. To confirm a regulatory effect of endocannabinoids on NK cell activation (9), in the present study NK cells were stimulated with anti-NK1.1 antibody obtaining a dose dependent inhibition of IFN-γ production by endocannabinoid 2-arachidonyl-glycerol (2-AG). Notably, adoptive transfer of CB2 -/- NK cells into WT hosts suppressed HDM-induced airway inflammation, suggesting that NK cells unrestrained by endogenous CB2 stimulation may act as gatekeepers to limit allergen responsiveness, eventually by mechanisms that include the regulation of allergy–promoting ILC2 cells.
Early studies in the seventies revealed a beneficial effect of inhaled or orally taken cannabinoids by dilating bronchial smooth muscles (12, 13). Since then, both anti- and pro–inflammatory effects of endocannabinoids have been reported, which has been attributed, in part, to bioactive metabolites and signalling by additional receptors, aside from CB1 and CB2, which are both expressed on human leukocytes (14). Several studies using mice lacking the CB2 receptor have shown exacerbated inflammation in different disease models (15). In contrast, Ferrini et al. (11) demonstrate in the HDM-induced asthma model using CB2 -/- mice that CB2 augments allergen responsiveness. Interestingly, an allergy promoting role of CB2 was also observed in an ear dermatitis model with CB2 -/- mice (16), and by the use of the CB2 receptor agonist JWH-133 in a OVA-induced asthma model with C57BL6 mice showing a direct effect of CB2 on eosinophil responses (17). This raises the question if the immunomodulatory role of CB2 may depend on the type of immune response (e.g. allergic versus non-allergic). Further studies will be required to address this question or to exclude potential confounding variables, e.g. related to the disease model or mouse strain used (18, 19). In this regard, abnormalities in the development of several lymphocyte subsets have been reported in CB2 -/- mice (20).
The finding by Ferrini et al. (11) that NK cells may control ILC2 cells in allergic responses is intriguing. The exact mechanisms remain to be explored, but may include effects of NK cell-derived cytokines, such as IFN–γ (21), or other divergent strategies that NK cells employ for the control of immune cells (22). Given that NK cells are implicated in immune responses to viruses, the findings by Ferrini et al. (11) might indicate a potential mechanistic link between viral infection and TH2-biased allergic airway disease (23). Furthermore, altered frequencies and functions of NK cells in asthma patients have previously been reported (24, 25). The study by Ferrini et al. (11) highlights the need to explore cellular networks including ILC cells in different individuals and phenotypes of allergic diseases to better understand these disorders at the clinical and mechanistic level (26).
References
1. von Gunten S, Cortinas-Elizondo F, Kollarik M, Beisswenger C, Lepper PM. Mechanisms and potential therapeutic targets in allergic inflammation: recent insights. Allergy 2013;68:1487-1498.
2. Muraro A, Fokkens WJ, Pietikainen S, Borrelli D, Agache I, Bousquet J, et al. European symposium on precision medicine in allergy and airways diseases: report of the European Union parliament symposium (October 14, 2015). Rhinology 2015;53:303-307.
3. Licona-Limon P, Kim LK, Palm NW, Flavell RA. TH2, allergy and group 2 innate lymphoid cells.
Nat Immunol 2013;14:536-542.
4. Spits H, Artis D, Colonna M, Diefenbach A, Di Santo JP, Eberl G, et al. Innate lymphoid cells–a proposal for uniform nomenclature. Nat Rev Immunol 2013;13:145-149.
5. Schneider C, Smith DF, Cummings RD, Boligan KF, Hamilton RG, Bochner BS, et al. The human IgG anti-carbohydrate repertoire exhibits a universal architecture and contains specificity for microbial attachment sites. Sci Transl Med 2015;7:269ra261.
6. Pashova S, Schneider C, von Gunten S, Pashov A. Antibody Repertoire Profiling with Mimotope Arrays. Hum Vaccin Immunother; in press.
7. Lanier LL. Up on the tightrope: natural killer cell activation and inhibition. Nat Immunol 2008;9:495-502.
8. Jandus C, Boligan KF, Chijioke O, Liu H, Dahlhaus M, Demoulins T, et al. Interactions between Siglec-7/9 receptors and ligands influence NK cell-dependent tumor immunosurveillance. J Clin Invest 2014;124:1810–1820.
9. Massi P, Fuzio D, Vigano D, Sacerdote P, Parolaro D. Relative involvement of cannabinoid CB(1) and CB(2) receptors in the Delta(9)-tetrahydrocannabinol-induced inhibition of natural killer
activity. Eur J Pharmacol 2000;387:343-347.
10. Miljkovic D, Bassiouni A, Cooksley C, Ou J, Hauben E, Wormald PJ, et al. Association between group 2 innate lymphoid cells enrichment, nasal polyps and allergy in chronic rhinosinusitis. Allergy
2014;69:1154-1161.
11. Ferrini ME, Hong S, Stierle A, Stierle D, Stella N, Roberts K, et al. CB2 receptors regulate natural killer cells that limit allergic airway inflammation in a murine model of asthma. Allergy 2017;ADD SAME ISSUE (EDITORIAL ON THIS ARTICLE).
12. Tashkin DP, Reiss S, Shapiro BJ, Calvarese B, Olsen JL, Lodge JW. Bronchial effects of aerosolized delta 9-tetrahydrocannabinol in healthy and asthmatic subjects. Am Rev Respir Dis 1977;115:57-65.
13. Abboud RT, Sanders HD. Effect of oral administration of delta-tetrahydrocannabinol on airway mechanics in normal and asthmatic subjects. Chest 1976;70:480-485.
14. Turcotte C, Chouinard F, Lefebvre JS, Flamand N. Regulation of inflammation by cannabinoids, the endocannabinoids 2-arachidonoyl-glycerol and arachidonoyl-ethanolamide, and
their metabolites. J Leukoc Biol 2015;97:1049-1070.
15. Turcotte C, Blanchet MR, Laviolette M, Flamand N. The CB2 receptor and its role as a regulator of inflammation. Cell Mol Life Sci 2016;73:4449-4470.
16. Mimura T, Oka S, Koshimoto H, Ueda Y, Watanabe Y, Sugiura T. Involvement of the endogenous cannabinoid 2 ligand 2-arachidonyl glycerol in allergic inflammation. Int Arch Allergy
Immunol 2012;159:149-156.
17. Frei RB, Luschnig P, Parzmair GP, Peinhaupt M, Schranz S, Fauland A, et al. Cannabinoid receptor 2 augments eosinophil responsiveness and aggravates allergen-induced pulmonary
inflammation in mice. Allergy 2016;71:944-956.
18. von Gunten S, Shoenfeld Y, Blank M, Branch DR, Vassilev T, Kasermann F, et al. IVIG pluripotency and the concept of Fc-sialylation: challenges to the scientist. Nat Rev Immunol 2014;14:349.
19. Quast I, Keller CW, Weber P, Schneider C, von Gunten S, Lunemann JD. Protection from experimental autoimmune encephalomyelitis by polyclonal IgG requires adjuvant-induced
inflammation. J Neuroinflammation 2016;13:42.
20. Ziring D, Wei B, Velazquez P, Schrage M, Buckley NE, Braun J. Formation of B and T cell subsets require the cannabinoid receptor CB2. Immunogenetics 2006;58:714-725.
21. Moro K, Kabata H, Tanabe M, Koga S, Takeno N, Mochizuki M, et al. Interferon and IL–27 antagonize the function of group 2 innate lymphoid cells and type 2 innate immune responses. Nat
Immunol 2016;17:76-86.
22. Crouse J, Xu HC, Lang PA, Oxenius A. NK cells regulating T cell responses: mechanisms and outcome. Trends Immunol 2015;36:49-58.
23. Lan F, Zhang N, Gevaert E, Zhang L, Bachert C. Viruses and bacteria in Th2-biased allergic airway disease. Allergy 2016;71:1381-1392.
24. Lin SJ, Chang LY, Yan DC, Huang YJ, Lin TJ, Lin TY. Decreased intercellular adhesion molecule–
1 (CD54) and L-selectin (CD62L) expression on peripheral blood natural killer cells in asthmatic children with acute exacerbation. Allergy 2003;58:67-71.
25. Jira M, Antosova E, Vondra V, Strejcek J, Mazakova H, Prazakova J. Natural killer and interleukin-2 induced cytotoxicity in asthmatics. I. Effect of acute antigen-specific challenge. Allergy
1988;43:294-298.
26. Bousquet J, Anto JM, Akdis M, Auffray C, Keil T, Momas I, et al. Paving the way of systems biology and precision medicine in allergic diseases: the MeDALL success story: Mechanisms of the
Development of ALLergy; EU FP7-CP-IP; Project No: 261357; 2010-2015. Allergy 2016;71:1513–1525.
Figures
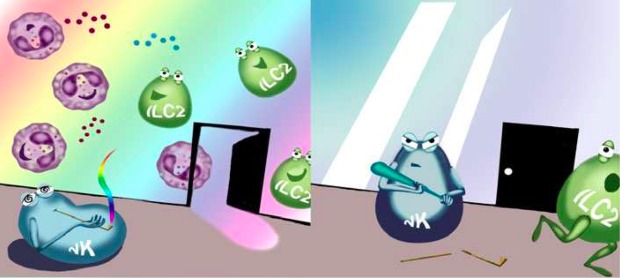
Figure 1 Putative role of NK cells as gatekeepers of the allergic party, which may involve the control of allergy-promoting ILC2 cells. The NK cell gatekeeper’s attention is impaired by signalling through the cannabinoid receptor CB2 (left panel), and enhanced in absence of CB2, represented as broken cannabis pipe in the cartoon (right panel). Illustration by Aldona von Gunten.
Accepted Article
This article is protected by copyright. All rights reserved.
