

![]()
Inhibition of Glioma Growth in Vivo by Selective Activation of the CB2 Cannabinoid Receptor 1
![]()
Inhibition of Glioma Growth in Vivo by Selective Activation of the CB2 Cannabinoid Receptor 1Cristina Sánchez2, María L. de Ceballos2, Teresa Gómez del Pulgar2, Daniel Rueda, César Corbacho, Guillermo Velasco, Ismael Galve-Roperh, John W. Huffman, Santiago Ramón y Cajal, and Manuel Guzmán3
+ Author Affiliations
Department of Biochemistry and Molecular Biology I, School of Biology, Complutense Un

+ Author Affiliations
Department of Biochemistry and Molecular Biology I, School of Biology, Complutense Un
iversity, 28040 Madrid, Spain [C. S., T. G. d. P., D. R., G. V., I. G-R., M. G.]; Neurodegeneration Group, Cajal Institute, CSIC, 28002 Madrid, Spain [M. L. d. C.]; Department of Pathology, Clínica Puerta de Hierro, 28035 Madrid, Spain [C. C., S. R. y C.]; and Department of Chemistry, Clemson University, Clemson, South Carolina 29634-1905 [J. W. H.]
Abstract
The development of new therapeutic strategies is essential for the management of gliomas, one of the most malignant forms of cancer. We have shown previously that the growth of the rat glioma C6 cell line is inhibited by psychoactive cannabinoids (I. Galve-Roperh et al., Nat. Med., 6: 313–319, 2000). These compounds act on the brain and some other organs through the widely expressed CB1 receptor. By contrast, the other cannabinoid receptor subtype, the CB2 receptor, shows a much more restricted distribution and is absent from normal brain. Here we show that local administration of the selective CB2 agonist JWH-133 at 50 μg/day to Rag-2−/− mice induced a considerable regression of malignant tumors generated by inoculation of C6 glioma cells. The selective involvement of the CB2 receptor in this action was evidenced by: (a) the prevention by the CB2 antagonist SR144528 but not the CB1 antagonist SR141716; (b) the down-regulation of the CB2 receptor but not the CB1 receptor in the tumors; and (c) the absence of typical CB1-mediated psychotropic side effects. Cannabinoid receptor expression was subsequently examined in biopsies from human astrocytomas. A full 70% (26 of 37) of the human astrocytomas analyzed expressed significant levels of cannabinoid receptors. Of interest, the extent of CB2 receptor expression was directly related with tumor malignancy. In addition, the growth of grade IV human astrocytoma cells in Rag-2−/− mice was completely blocked by JWH-133 administration at 50 μg/day. Experiments carried out with C6 glioma cells in culture evidenced the internalization of the CB2 but not the CB1 receptor upon JWH-133 challenge and showed that selective activation of the CB2 receptor signaled apoptosis via enhanced ceramide synthesis de novo. These results support a therapeutic approach for the treatment of malignant gliomas devoid of psychotropic side effects.
INTRODUCTION
Cannabinoids, the active components of Cannabis sativa and their derivatives, exert a wide spectrum of central and peripheral actions, such as analgesia, anticonvulsion, anti-inflammation, and alleviation of both intraocular pressure and emesis. These effects are mediated by the activation of specific G protein-coupled receptors (1 , 2) . To date, two different cannabinoid receptors have been characterized and cloned from mammalian tissues: CB1 (3) and CB2 (4) . The central and most of the peripheral effects of cannabinoids rely on CB1 receptor activation. This receptor is found in high levels in the central nervous system, where it mediates cannabinoid psychoactivity, and is also present in peripheral nerve terminals, as well as in extra-neural sites, such as testis, uterus, vascular endothelium, eye, spleen, and tonsils (1, 2, 3, 4, 5, 6) . By contrast, the CB2 receptor is believed to be solely expressed in cells and organs of the immune system and is unrelated to cannabinoid psychoactivity (1 , 2) . The discovery of a family of endogenous ligands of cannabinoid receptors, the so-called endocannabinoids (7, 8, 9) , together with their specific mechanisms of synthesis and inactivation (10 , 11) , have focused much attention on cannabinoids during the last few years.
Marijuana and its derivatives have been used in medicine for many centuries, and nowadays, there is a renaissance in the study of the therapeutic effects of cannabinoids, which constitutes a widely debated issue with ample scientific and social relevance. Ongoing research is determining whether cannabinoid ligands may be effective agents in the treatment of pain (12 , 13) , glaucoma (14) , neurodegenerative disorders such as Parkinson’s disease (15) and multiple sclerosis (16) , and the wasting and emesis associated with AIDS and cancer chemotherapy (14) . In addition, cannabinoids might be potential antitumoral agents because of their ability to inhibit the growth of various types of cancer cells in culture (17, 18, 19) . Moreover, in laboratory animals, cannabinoids induce the regression of gliomas, one of the most malignant forms of cancer whose current treatment in patients is usually ineffective or just palliative (20) . This growth-inhibiting effect was exerted by two psychoactive cannabinoids, namely THC, 4 the main active component of marijuana, and WIN-55,212-2, a nonselective synthetic cannabinoid agonist, pointing to the involvement of cannabinoid receptors (20) . It would be desirable, however, that cannabinoid-based therapeutic strategies were devoid of typical CB1 receptor-mediated psychotropic side effects. Hence, the recent synthesis of selective CB2 agonists (21 , 22) opens a very attractive clinical possibility. The present work was therefore undertaken to test: (a) if gliomas, including those of human origin, express functional CB2 receptors; (b) if selective CB2 receptor activation exerts an antitumoral action in vivo; and (c) what may be the mechanism of that potential CB2-mediated antitumoral action.
MATERIALS AND METHODS
Materials.
JWH-133 was prepared in Dr. J. W. Huffman’s laboratory (22) . SR141716 and SR144528 were kindly given by Sanofi Recherche (Montpellier, France). The anti-CB1 receptor antibody (raised against residues 1–14 of the rat CB1 receptor) was kindly given by Dr. A. Howlett (North Carolina Central University, Durham, NC). The anti-CB2 receptor antibody (raised against residues 20–33 of the human CB2 receptor) was from Cayman Chemicals (Ann Arbor, MI). The Cy3-conjugated antirabbit IgG was from Amersham-Pharmacia (Buckinghamshire, United Kingdom). WIN-55,212-2 was from Sigma Chemical Co. (St. Louis, MO).
Glioma Cell Culture and Death.
The rat glioma C6 line was cultured as described before (18) . Cell viability was determined by the 3-4,5-dimethylthiazol-2,5-
Antitumoral Action of Cannabinoids in Vivo.
Tumors were induced in mice deficient in recombination activating gene 2 (Rag-2−/−), which lacks mature T and B cells (23) , by s.c. flank inoculation of 5 × 106 tumor cells (either C6 glioma cells or human astrocytoma cells) in PBS supplemented with 0.1% glucose. When tumors had reached an average volume of 250 mm3 (range, 200–300 mm3), animals were assigned randomly to various groups and injected intratumorally ≤8 (C6 glioma cells) or 25 (human astrocytoma cells) days with vehicle or 50 μg of cannabinoid ligand (JWH-133, WIN-55,212-2, SR141716, and/or SR144528) per day in 100 μl of PBS supplemented with 5 mg/ml defatted and dialyzed BSA. Tumors were measured with external caliper, and volume was calculated as (4π/3) × (width/2)2 × (length/2).
Motor Activity.
Motor activity (ambulation, rearing, and time of inactivity) was tested after intratumoral injection to C6-cell glioma-bearing mice, exactly under the aforementioned conditions, with vehicle or 50 μg of cannabinoid agonist (JWH-133 or WIN-55,212-2) in an open field (30 × 30 cm, divided into 16 squares of equal size) for 15 min. Animals were not habituated previously to the open field.
Immunofluorescence Analysis of Cannabinoid Receptors.
After cannabinoid treatment, cells were washed with PBS and fixed in cold acetone for 5 min. Immunolabeling was performed according to Hsieh et al. (24) . Cells were incubated with the anti-CB1 or anti-CB2 receptor antibodies (1:500) in the latter buffer for 3–4 h at room temperature and overnight at 4°C in a humid chamber. After washing with PBS, cultures were further incubated for 90 min with a Cy3-conjugated antirabbit IgG (1:800), washed first with PBS and then with 50 mM Tris-HCl (pH 7.4), and mounted with 50% glycerol. Preparations were analyzed with a Zeiss confocal laser-scanning microscope (excitation 550 nm, emission 565 nm). There was no labeling when the primary antibody was omitted (data not shown). In other experiments, 40-μm cryostat sections from C6 glioma-cell tumors were similarly treated after fixation with 4% paraformaldehyde in 0.1 M sodium phosphate buffer (pH 7.4) for 30 min. Finally, immunostaining was also performed in 5-μm sections of human astrocytomas (from the files of the Department of Pathology of Clínica Puerta de Hierro and with informed consent from each subject) pre-embedded in paraffin after deparaffination of the sections. Deparaffinized sections were incubated in DAKO sodium citrate buffer in a pressure cooker for 4 min, and immunohistochemical staining with the anti-CB1 and CB2 receptor antibodies was evaluated using the avidin-biotin/peroxidase technique in a Horizon Dako (Mesip program) automated immunohistochemical stainer, according to the manufacturer instructions.
Western Blot Analysis of Cannabinoid Receptors.
Particulate cell or tumor fractions were subjected to SDS-PAGE, and proteins were transferred from the gels onto polyvinylidene fluoride membranes. The blots were incubated with the aforementioned antibodies against the CB1 receptor (1:5000) or the CB2 receptor (1:2000). Samples were finally subjected to luminography with an enhanced chemiluminescence detection kit (20) .
SPT Activity.
SPT activity was determined in digitonin-permeabilized C6 glioma cells as the incorporation of radiolabeled L-serine into ketosphinganine by a new procedure (25) . Briefly, the medium was aspirated, and cells were washed twice with PBS. Reactions were started by the addition of 100 mM HEPES (pH 8.3), 200 mM sucrose, 2.5 mM EDTA, 5 mM dithioerythritol, 50 μM pyridoxal phosphate, 1 mg/ml BSA, 70 μg/ml digitonin, 0.3 mM palmitoyl-CoA, and 0.25 mM L-[U-14C]serine (3 μCi/assay). After 45 min, reactions were stopped with 0.5 M NH4OH, and [14C]ketosphinganine product was extracted with chloroform/methanol/1% NaCl.
ERK Activity.
Cells were washed and lysed, supernatants were obtained, and ERK activity was determined as the incorporation of [γ-32P]ATP into a specific peptide substrate (20) .
Statistics.
Results shown represent means ± SD. Statistical analysis of cannabinoid receptor expression (Table 1) ⇓ was performed by the χ2 test. For the rest of the data, ANOVA, with a post hoc analysis by the Student-Neuman-Keuls test, was used.
View this table: http://
Table 1
Cannabinoid receptor expression in human astrocytomas of different malignancya
RESULTS
Regression of C6-Cell Gliomas in Vivo by Selective CB2 Receptor Activation.
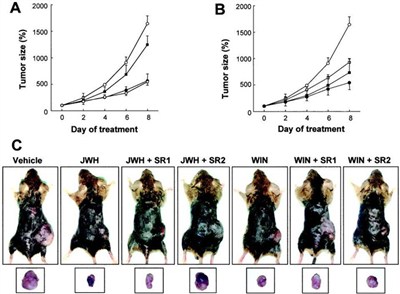
Cannabinoid-based therapeutic strategies should be as devoid as possible of psychotropic side effects. Because cannabinoid psychoactivity is mediated by the CB1 receptor (1 , 2) and C6 glioma cells in culture express the CB2 receptor protein (20) , we evaluated if selective CB2 receptor activation inhibits the growth of malignant gliomas in vivo by using the selective CB2 agonist JWH-133 (Ki = 677 nM for CB1 and 3.4 nM for CB2; Ref. 22 ). WIN-55,212-2 (Ki = 1.9 nM for CB1 and 0.3 nM for CB2) was used as a reference for nonselective cannabinoid receptor activation (26) . As shown in Fig. 1A ⇓ , tumor growth was remarkably lower in mice intratumorally injected with 50 μg/day of JWH-133 than in control animals. The magnitude of this antiproliferative action was comparable with that exerted by WIN-55,212-2 at equal doses (Fig. 1B) ⇓ .
Fig. 1. http://
Regression of C6-cell gliomas in vivo upon selective CB2 receptor activation. C6 glioma cells were injected s.c. in mice. When tumors had reached the desired size (day 0), animals were treated with either vehicle (?), cannabinoid alone (•), cannabinoid plus SR141716 (?), or cannabinoid plus SR144528 (?) for 8 days (n = 6 for each experimental group). A, JWH-133 administration. B, WIN-55,212-2 administration. C, examples of s.c. gliomas in the flank of mice (top panel) and after dissection (bottom panel) after the indicated treatments for 8 days. JWH, JWH-133; WIN, WIN-55,212-2; SR1, SR141716; SR2, SR144528.
To evaluate the possible implication of the CB1 and CB2 cannabinoid receptors in the antitumoral action of JWH-133 in vivo, the effect of selective cannabinoid receptor antagonists was evaluated. The selective CB2 antagonist SR144528 (Ki = 437 nM for CB1 and 0.6 nM for CB2; Ref. 27 ), but not the selective CB1 antagonist SR141716 (Ki = 5.6 nM for CB1 and >1 μM for CB2; Ref. 28 ), prevented JWH-133-induced tumor regression (Fig. 1A) ⇓ . By contrast, the antitumoral effect of WIN-55,212-2 was largely evident upon coadministration of any of the two antagonists (Fig. 1B) ⇓ . Neither SR141716 nor SR144528 per se exerted a significant effect on tumor growth (data not shown). Examples of tumor-bearing mice and of dissected tumors after cannabinoid treatment for 8 days are shown in Fig. 1C ⇓ .
Under the conditions in which it induced tumor regression, JWH-133 administration led to no significant alteration of typical CB1-mediated behavioral parameters, such as ambulation, rearing, and time of inactivity in an open field trial. By contrast, although cannabinoids were inoculated locally at the site of the tumor, WIN-55,212-2 treatment produced a clear inhibition of those parameters. Thus, ambulation (in number of squares crossed) was 124 ± 21 (vehicle), 131 ± 26 (JWH-133), and 89 ± 35 (WIN-55,212-2); rearing (in number of rears) was 16 ± 3 (vehicle), 22 ± 11 (JWH-133), and 5 ± 5 (WIN-55,212-2); and time of inactivity (in s) was 6 ± 6 (vehicle), 4 ± 3 (JWH-133), and 204 ± 95 (WIN-55,212-2; n = 6 for each condition).
Expression and Dynamics of Cannabinoid Receptors in C6 Glioma Cells in Culture and in Vivo.
The presence and dynamics of CB1 and CB2 receptors in C6 glioma cells were examined by immunofluorescence experiments. As shown in Fig. 2a ⇓ (A), a quite homogeneous signal in the plasma membrane and the cytoplasm was detected for both receptors in cultured cells. Exposure to cannabinoid agonists is known to induce internalization of cannabinoid receptors (1 , 2 , 24) . Likewise, upon WIN-55,212-2 exposure, CB1 and CB2 immunostaining, although still observed in the cytoplasm, turned to be more intense in the perinuclear region (Fig. 2a ⇓ , A). By contrast, JWH-133 action was only evident on the CB2 receptor, CB1 signal remaining unaffected by this cannabinoid (Fig. 2a ⇓ , A). Cannabinoid receptor immunoreactivity was also detected in C6-cell gliomas obtained from tumor-bearing mice. CB1 receptor expression was not significantly affected by JWH-133 and SR144528 administration, but in SR141716-treated tumors, a slight increase in the labeling was noted (Fig. 2a ⇓ , B). By contrast, CB2 receptor expression was reduced by JWH-133, whereas SR144528 inoculation significantly increased it and blocked the JWH-133 effect. SR141716 did not significantly affect the labeling (Fig. 2a ⇓ , B). These observations are in agreement with the well-known tolerance that occurs after chronic cannabinoid administration (1 , 2) and further support the notion that CB2 receptors are functional in C6 glioma cells, JWH-133 being a highly selective CB2 agonist in our experimental system.
Fig. 2. http://
a. Immunofluorescence analysis of cannabinoid receptor dynamics in cultured glioma cells and in gliomas. Images of representative experiments are shown. Similar results were obtained in three other experiments for each experimental condition. In A, C6 glioma cells were cultured for 30 min in the absence or presence of 100 nM JWH-133 or WIN-55,212-2. B, C6-cell gliomas. Mice were treated as in Fig. 1 ⇓ for 8 days, tumors were dissected, and immunomicroscopy analysis was performed. Abbreviations as in Fig. 1 ⇓ . Fig. 2b ⇓ . Regression of human astrocytomas in vivo upon selective CB2 receptor activation. A, example of CB2 receptor expression in a grade IV human astrocytoma (HA) as assessed by immunomicroscopy analysis of a tumor section and of cells derived from the tumor. Western blot analysis of the tumor was also performed using C6 glioma cells as a control. In B, cells from the tumor shown in A were injected s.c. in mice. When tumors had reached the desired size (day 0), animals were treated with either vehicle (?) or JWH-133 (•) for 25 days (n = 6 for each experimental group). C, examples of s.c. gliomas in the flank of mice (top panel) and after dissection (bottom panel) after treatment with vehicle or JWH-133 (JWH) for 25 days.
Regression of Human Astrocytomas in Vivo by Selective CB2 Receptor Activation.
The presence of CB1 and CB2 receptors was analyzed by immunohistochemistry in biopsies from human astrocytomas of low (grades I–II) and high (grades III–IV) malignancy. High malignancy tumors were diagnosed in all cases as pure astrocytomas, whereas some of the low malignancy tumors were mixed oligoastrocytomas. However, no differences were observed in the distribution of cannabinoid receptors between pure and mixed tumors (data not shown). A full 70% (26 of 37) of the total tumors analyzed expressed measurable levels of cannabinoid receptors (i.e., CB1 and/or CB2). Among them, a similar percentage of low malignancy (4 of 12) and high malignancy tumors (6 of 25) expressed CB1 and CB2 receptors simultaneously. Of interest, the extent of CB2 receptor expression was related with tumor malignancy. Thus, as shown in Table 1 ⇓ , the proportion of grade IV astrocytomas showing high/very high CB2 immunoreactivity doubled that of grades I–III tumors. Moreover, CB2 receptor expression was markedly enhanced in grade IV astrocytomas compared with CB1 receptor expression.
Given the inhibition of C6-cell glioma growth by selective CB2 receptor activation (Fig. 1) ⇓ , we evaluated the effect of JWH-133 treatment on the growth of highly malignant (grade IV) human astrocytoma cells in vivo. Immunofluorescence microscopy and Western blot analyses evidenced the expression of the CB2 receptor in the inoculated cells (Fig. 2b ⇓ , A). This particular tumor also expressed the CB1 receptor (data not shown). Of interest, JWH-133 administration completely blocked the proliferation of the human astrocytoma (Fig. 2b ⇓ , B). Examples of tumor-bearing mice and of dissected tumors after cannabinoid treatment for 25 days are shown in Fig. 2b ⇓ , C.
Selective CB2 Receptor Activation Signals Apoptosis of C6 Glioma Cells via Ceramide Synthesis de Novo.
We have shown previously that THC-induced apoptosis of C6 glioma cells relies on the sustained generation of the proapoptotic lipid ceramide (20 , 29) . To obtain further evidence for the specificity of the JWH-133 antitumoral action, experiments were carried out with L-cycloserine, a selective competitive inhibitor of SPT, the enzyme which catalyzes the pace-setting step of ceramide synthesis de novo (30) . As shown in Fig. 3, A and B ⇓ , exposure to JWH-133 induced apoptosis of C6 glioma cells, and this effect was prevented by L-cycloserine. Moreover, L-cycloserine was able to suppress JWH-133-evoked SPT induction (Fig. 3C) ⇓ . We also tested the effect of L-cycloserine on JWH-133-induced ERK activation, as ERK seems to be the downstream target of ceramide in THC-evoked apoptosis of C6 glioma cells (20 , 29) . Thus, blockade of ceramide synthesis de novo with L-cycloserine abrogated JWH-133-induced ERK activation (Fig. 3D) ⇓ , indicating that JWH-133, like THC, signals apoptosis via ceramide synthesis de novo and ERK activation.
Fig. 3. http://
Selective CB2 receptor activation induces apoptosis of C6 glioma cells via ceramide synthesis de novo. C6 glioma cells were cultured in the absence or presence of 100 nM JWH-133 (JWH) and/or 0.5 mM L-cycloserine (CS) for 5 days. A, TUNEL staining and phase-contrast micrographs. Images of a representative experiment are shown. Similar results were obtained in two other experiments. B, cell viability (n = 6). C, SPT activity (n = 4). D, ERK activity (n = 6). ∗, significantly different (P < 0.01) from incubations with no additions.
DISCUSSION
Recent studies have dealt with the antiproliferative effect of cannabinoids on different transformed cells. However, this property of cannabis compounds was first reported 25 years ago by Munson et al. (31) , who showed that cannabinoids may inhibit the growth of Lewis lung adenocarcinoma, B-tropic Friend leukemia virus-induced splenomegaly, and L1210 leukemia cells in vivo. Although these observations were actually promising, further investigations were not performed on this area until a few years ago. Nowadays, there are data demonstrating that cannabinoids inhibit the growth of transformed neural, breast, and prostate cells in culture (32) and induce the regression of gliomas in vivo (Ref. 20 and the present study). The case of gliomas is of particular interest because they are one of the most malignant forms of cancer, resulting in the death of affected patients within months after diagnosis. Conventional therapies, including surgery, radiotherapy, chemotherapy, and immunotherapy, are usually ineffective or just palliative (33 , 34) . The most recent strategies for glioma treatment are focused on gene therapy, but no trial performed thus far has been significantly successful (33, 34, 35) . It is therefore essential to develop new therapeutic strategies for the management of gliomas and most likely to combine some of them to obtain significant clinical results. One of these alternative therapeutic approaches might be on the basis of the use of nonpsychoactive cannabinoid ligands, because these compounds induce apoptosis in vitro and inhibition of tumor growth without significant collateral effects in vivo. Because of the current inability to kill target glioma cells with sufficient efficacy to obtain significant tumor regression in the clinical practice, the combined use of drugs that inhibit tumor growth with cytotoxic approaches may be envisaged as the most feasible anticancer strategy in the near future. Of interest, cannabinoid apoptotic action relies on the generation of ceramide, a lipid second messenger that may have antitumoral properties either alone (36) or in combined therapies (37) .
We are nevertheless aware that under certain circumstances, cannabinoids may be immunosuppressive compounds by acting on immune organs and cells via CB2 receptors, and this would be expected to inhibit host antitumor immunity. As a matter of fact, Zhu et al. (38) have recently reported that i.p. THC injection to immune-competent mice for 4–6 weeks leads to an accelerated growth of tumor implants in two different murine lung cancer models. This effect, although not evidenced in the former report by Munson et al. (31) , was shown to rely on the CB2-dependent inhibition of the capacity of antigen-presenting cells and T cells to generate alloreactivity (38) . It is therefore possible that cannabinoids exert a dual effect on tumor growth, i.e., a direct antiproliferative effect (Ref. 20 and the present study) and an indirect growth-enhancing effect via inhibition of immunogenicity (38) . Factors such as the route of drug administration (local versus systemic), the timing of drug delivery (short-term versus long-term treatment), and the intrinsic capacity of a particular tumor cell to respond to cannabinoids (e.g., presence versus absence of cannabinoid receptors) might determine the balance between tumor progression and regression. In any event, the present study, together with our previous observations (20) , shows that the antitumoral action of cannabinoids on gliomas may be exerted either via the CB1 receptor or via the CB2 receptor. The attractive possibility of finding cannabinoid-based therapeutic strategies for neural diseases devoid of nondesired CB1-mediated psychotropic side effects is also opened by the possible implication of the CB2 receptor in the control of pain initiation (12) and multiple sclerosis-linked spasticity (16) . Moreover, our data support the notion that the CB2 receptor might serve as a diagnostic marker of glial cell proliferation/malignancy, in line with what Valk et al. (39 , 40) have reported for myeloid cell growth and transformation during leukemogenesis.

Acknowledgments
We thank J. L. Gil, M. E. Fernández de Molina, C. Bailón, C. Hernández, and J. Palacín for their expert technical assistance. This article is dedicated to Dr. Math J. H. Geelen, outstanding on personal and scientific grounds, on his retirement as Professor of Veterinary Biochemistry.
Abstract
The development of new therapeutic strategies is essential for the management of gliomas, one of the most malignant forms of cancer. We have shown previously that the growth of the rat glioma C6 cell line is inhibited by psychoactive cannabinoids (I. Galve-Roperh et al., Nat. Med., 6: 313–319, 2000). These compounds act on the brain and some other organs through the widely expressed CB1 receptor. By contrast, the other cannabinoid receptor subtype, the CB2 receptor, shows a much more restricted distribution and is absent from normal brain. Here we show that local administration of the selective CB2 agonist JWH-133 at 50 μg/day to Rag-2−/− mice induced a considerable regression of malignant tumors generated by inoculation of C6 glioma cells. The selective involvement of the CB2 receptor in this action was evidenced by: (a) the prevention by the CB2 antagonist SR144528 but not the CB1 antagonist SR141716; (b) the down-regulation of the CB2 receptor but not the CB1 receptor in the tumors; and (c) the absence of typical CB1-mediated psychotropic side effects. Cannabinoid receptor expression was subsequently examined in biopsies from human astrocytomas. A full 70% (26 of 37) of the human astrocytomas analyzed expressed significant levels of cannabinoid receptors. Of interest, the extent of CB2 receptor expression was directly related with tumor malignancy. In addition, the growth of grade IV human astrocytoma cells in Rag-2−/− mice was completely blocked by JWH-133 administration at 50 μg/day. Experiments carried out with C6 glioma cells in culture evidenced the internalization of the CB2 but not the CB1 receptor upon JWH-133 challenge and showed that selective activation of the CB2 receptor signaled apoptosis via enhanced ceramide synthesis de novo. These results support a therapeutic approach for the treatment of malignant gliomas devoid of psychotropic side effects.
INTRODUCTION
Cannabinoids, the active components of Cannabis sativa and their derivatives, exert a wide spectrum of central and peripheral actions, such as analgesia, anticonvulsion, anti-inflammation, and alleviation of both intraocular pressure and emesis. These effects are mediated by the activation of specific G protein-coupled receptors (1 , 2) . To date, two different cannabinoid receptors have been characterized and cloned from mammalian tissues: CB1 (3) and CB2 (4) . The central and most of the peripheral effects of cannabinoids rely on CB1 receptor activation. This receptor is found in high levels in the central nervous system, where it mediates cannabinoid psychoactivity, and is also present in peripheral nerve terminals, as well as in extra-neural sites, such as testis, uterus, vascular endothelium, eye, spleen, and tonsils (1, 2, 3, 4, 5, 6) . By contrast, the CB2 receptor is believed to be solely expressed in cells and organs of the immune system and is unrelated to cannabinoid psychoactivity (1 , 2) . The discovery of a family of endogenous ligands of cannabinoid receptors, the so-called endocannabinoids (7, 8, 9) , together with their specific mechanisms of synthesis and inactivation (10 , 11) , have focused much attention on cannabinoids during the last few years.
Marijuana and its derivatives have been used in medicine for many centuries, and nowadays, there is a renaissance in the study of the therapeutic effects of cannabinoids, which constitutes a widely debated issue with ample scientific and social relevance. Ongoing research is determining whether cannabinoid ligands may be effective agents in the treatment of pain (12 , 13) , glaucoma (14) , neurodegenerative disorders such as Parkinson’s disease (15) and multiple sclerosis (16) , and the wasting and emesis associated with AIDS and cancer chemotherapy (14) . In addition, cannabinoids might be potential antitumoral agents because of their ability to inhibit the growth of various types of cancer cells in culture (17, 18, 19) . Moreover, in laboratory animals, cannabinoids induce the regression of gliomas, one of the most malignant forms of cancer whose current treatment in patients is usually ineffective or just palliative (20) . This growth-inhibiting effect was exerted by two psychoactive cannabinoids, namely THC, 4 the main active component of marijuana, and WIN-55,212-2, a nonselective synthetic cannabinoid agonist, pointing to the involvement of cannabinoid receptors (20) . It would be desirable, however, that cannabinoid-based therapeutic strategies were devoid of typical CB1 receptor-mediated psychotropic side effects. Hence, the recent synthesis of selective CB2 agonists (21 , 22) opens a very attractive clinical possibility. The present work was therefore undertaken to test: (a) if gliomas, including those of human origin, express functional CB2 receptors; (b) if selective CB2 receptor activation exerts an antitumoral action in vivo; and (c) what may be the mechanism of that potential CB2-mediated antitumoral action.
MATERIALS AND METHODS
Materials.
JWH-133 was prepared in Dr. J. W. Huffman’s laboratory (22) . SR141716 and SR144528 were kindly given by Sanofi Recherche (Montpellier, France). The anti-CB1 receptor antibody (raised against residues 1–14 of the rat CB1 receptor) was kindly given by Dr. A. Howlett (North Carolina Central University, Durham, NC). The anti-CB2 receptor antibody (raised against residues 20–33 of the human CB2 receptor) was from Cayman Chemicals (Ann Arbor, MI). The Cy3-conjugated antirabbit IgG was from Amersham-Pharmacia (Buckinghamshire, United Kingdom). WIN-55,212-2 was from Sigma Chemical Co. (St. Louis, MO).
Glioma Cell Culture and Death.
The rat glioma C6 line was cultured as described before (18) . Cell viability was determined by the 3-4,5-dimethylthiazol-2,5-
Antitumoral Action of Cannabinoids in Vivo.
Tumors were induced in mice deficient in recombination activating gene 2 (Rag-2−/−), which lacks mature T and B cells (23) , by s.c. flank inoculation of 5 × 106 tumor cells (either C6 glioma cells or human astrocytoma cells) in PBS supplemented with 0.1% glucose. When tumors had reached an average volume of 250 mm3 (range, 200–300 mm3), animals were assigned randomly to various groups and injected intratumorally ≤8 (C6 glioma cells) or 25 (human astrocytoma cells) days with vehicle or 50 μg of cannabinoid ligand (JWH-133, WIN-55,212-2, SR141716, and/or SR144528) per day in 100 μl of PBS supplemented with 5 mg/ml defatted and dialyzed BSA. Tumors were measured with external caliper, and volume was calculated as (4π/3) × (width/2)2 × (length/2).
Motor Activity.
Motor activity (ambulation, rearing, and time of inactivity) was tested after intratumoral injection to C6-cell glioma-bearing mice, exactly under the aforementioned conditions, with vehicle or 50 μg of cannabinoid agonist (JWH-133 or WIN-55,212-2) in an open field (30 × 30 cm, divided into 16 squares of equal size) for 15 min. Animals were not habituated previously to the open field.
Immunofluorescence Analysis of Cannabinoid Receptors.
After cannabinoid treatment, cells were washed with PBS and fixed in cold acetone for 5 min. Immunolabeling was performed according to Hsieh et al. (24) . Cells were incubated with the anti-CB1 or anti-CB2 receptor antibodies (1:500) in the latter buffer for 3–4 h at room temperature and overnight at 4°C in a humid chamber. After washing with PBS, cultures were further incubated for 90 min with a Cy3-conjugated antirabbit IgG (1:800), washed first with PBS and then with 50 mM Tris-HCl (pH 7.4), and mounted with 50% glycerol. Preparations were analyzed with a Zeiss confocal laser-scanning microscope (excitation 550 nm, emission 565 nm). There was no labeling when the primary antibody was omitted (data not shown). In other experiments, 40-μm cryostat sections from C6 glioma-cell tumors were similarly treated after fixation with 4% paraformaldehyde in 0.1 M sodium phosphate buffer (pH 7.4) for 30 min. Finally, immunostaining was also performed in 5-μm sections of human astrocytomas (from the files of the Department of Pathology of Clínica Puerta de Hierro and with informed consent from each subject) pre-embedded in paraffin after deparaffination of the sections. Deparaffinized sections were incubated in DAKO sodium citrate buffer in a pressure cooker for 4 min, and immunohistochemical staining with the anti-CB1 and CB2 receptor antibodies was evaluated using the avidin-biotin/peroxidase technique in a Horizon Dako (Mesip program) automated immunohistochemical stainer, according to the manufacturer instructions.
Western Blot Analysis of Cannabinoid Receptors.
Particulate cell or tumor fractions were subjected to SDS-PAGE, and proteins were transferred from the gels onto polyvinylidene fluoride membranes. The blots were incubated with the aforementioned antibodies against the CB1 receptor (1:5000) or the CB2 receptor (1:2000). Samples were finally subjected to luminography with an enhanced chemiluminescence detection kit (20) .
SPT Activity.
SPT activity was determined in digitonin-permeabilized C6 glioma cells as the incorporation of radiolabeled L-serine into ketosphinganine by a new procedure (25) . Briefly, the medium was aspirated, and cells were washed twice with PBS. Reactions were started by the addition of 100 mM HEPES (pH 8.3), 200 mM sucrose, 2.5 mM EDTA, 5 mM dithioerythritol, 50 μM pyridoxal phosphate, 1 mg/ml BSA, 70 μg/ml digitonin, 0.3 mM palmitoyl-CoA, and 0.25 mM L-[U-14C]serine (3 μCi/assay). After 45 min, reactions were stopped with 0.5 M NH4OH, and [14C]ketosphinganine product was extracted with chloroform/methanol/1% NaCl.
ERK Activity.
Cells were washed and lysed, supernatants were obtained, and ERK activity was determined as the incorporation of [γ-32P]ATP into a specific peptide substrate (20) .
Statistics.
Results shown represent means ± SD. Statistical analysis of cannabinoid receptor expression (Table 1) ⇓ was performed by the χ2 test. For the rest of the data, ANOVA, with a post hoc analysis by the Student-Neuman-Keuls test, was used.
View this table: http://
Table 1
Cannabinoid receptor expression in human astrocytomas of different malignancya
RESULTS
Regression of C6-Cell Gliomas in Vivo by Selective CB2 Receptor Activation.
Cannabinoid-based therapeutic strategies should be as devoid as possible of psychotropic side effects. Because cannabinoid psychoactivity is mediated by the CB1 receptor (1 , 2) and C6 glioma cells in culture express the CB2 receptor protein (20) , we evaluated if selective CB2 receptor activation inhibits the growth of malignant gliomas in vivo by using the selective CB2 agonist JWH-133 (Ki = 677 nM for CB1 and 3.4 nM for CB2; Ref. 22 ). WIN-55,212-2 (Ki = 1.9 nM for CB1 and 0.3 nM for CB2) was used as a reference for nonselective cannabinoid receptor activation (26) . As shown in Fig. 1A ⇓ , tumor growth was remarkably lower in mice intratumorally injected with 50 μg/day of JWH-133 than in control animals. The magnitude of this antiproliferative action was comparable with that exerted by WIN-55,212-2 at equal doses (Fig. 1B) ⇓ .
Fig. 1. http://
Regression of C6-cell gliomas in vivo upon selective CB2 receptor activation. C6 glioma cells were injected s.c. in mice. When tumors had reached the desired size (day 0), animals were treated with either vehicle (?), cannabinoid alone (•), cannabinoid plus SR141716 (?), or cannabinoid plus SR144528 (?) for 8 days (n = 6 for each experimental group). A, JWH-133 administration. B, WIN-55,212-2 administration. C, examples of s.c. gliomas in the flank of mice (top panel) and after dissection (bottom panel) after the indicated treatments for 8 days. JWH, JWH-133; WIN, WIN-55,212-2; SR1, SR141716; SR2, SR144528.
To evaluate the possible implication of the CB1 and CB2 cannabinoid receptors in the antitumoral action of JWH-133 in vivo, the effect of selective cannabinoid receptor antagonists was evaluated. The selective CB2 antagonist SR144528 (Ki = 437 nM for CB1 and 0.6 nM for CB2; Ref. 27 ), but not the selective CB1 antagonist SR141716 (Ki = 5.6 nM for CB1 and >1 μM for CB2; Ref. 28 ), prevented JWH-133-induced tumor regression (Fig. 1A) ⇓ . By contrast, the antitumoral effect of WIN-55,212-2 was largely evident upon coadministration of any of the two antagonists (Fig. 1B) ⇓ . Neither SR141716 nor SR144528 per se exerted a significant effect on tumor growth (data not shown). Examples of tumor-bearing mice and of dissected tumors after cannabinoid treatment for 8 days are shown in Fig. 1C ⇓ .
Under the conditions in which it induced tumor regression, JWH-133 administration led to no significant alteration of typical CB1-mediated behavioral parameters, such as ambulation, rearing, and time of inactivity in an open field trial. By contrast, although cannabinoids were inoculated locally at the site of the tumor, WIN-55,212-2 treatment produced a clear inhibition of those parameters. Thus, ambulation (in number of squares crossed) was 124 ± 21 (vehicle), 131 ± 26 (JWH-133), and 89 ± 35 (WIN-55,212-2); rearing (in number of rears) was 16 ± 3 (vehicle), 22 ± 11 (JWH-133), and 5 ± 5 (WIN-55,212-2); and time of inactivity (in s) was 6 ± 6 (vehicle), 4 ± 3 (JWH-133), and 204 ± 95 (WIN-55,212-2; n = 6 for each condition).
Expression and Dynamics of Cannabinoid Receptors in C6 Glioma Cells in Culture and in Vivo.
The presence and dynamics of CB1 and CB2 receptors in C6 glioma cells were examined by immunofluorescence experiments. As shown in Fig. 2a ⇓ (A), a quite homogeneous signal in the plasma membrane and the cytoplasm was detected for both receptors in cultured cells. Exposure to cannabinoid agonists is known to induce internalization of cannabinoid receptors (1 , 2 , 24) . Likewise, upon WIN-55,212-2 exposure, CB1 and CB2 immunostaining, although still observed in the cytoplasm, turned to be more intense in the perinuclear region (Fig. 2a ⇓ , A). By contrast, JWH-133 action was only evident on the CB2 receptor, CB1 signal remaining unaffected by this cannabinoid (Fig. 2a ⇓ , A). Cannabinoid receptor immunoreactivity was also detected in C6-cell gliomas obtained from tumor-bearing mice. CB1 receptor expression was not significantly affected by JWH-133 and SR144528 administration, but in SR141716-treated tumors, a slight increase in the labeling was noted (Fig. 2a ⇓ , B). By contrast, CB2 receptor expression was reduced by JWH-133, whereas SR144528 inoculation significantly increased it and blocked the JWH-133 effect. SR141716 did not significantly affect the labeling (Fig. 2a ⇓ , B). These observations are in agreement with the well-known tolerance that occurs after chronic cannabinoid administration (1 , 2) and further support the notion that CB2 receptors are functional in C6 glioma cells, JWH-133 being a highly selective CB2 agonist in our experimental system.
Fig. 2. http://
a. Immunofluorescence analysis of cannabinoid receptor dynamics in cultured glioma cells and in gliomas. Images of representative experiments are shown. Similar results were obtained in three other experiments for each experimental condition. In A, C6 glioma cells were cultured for 30 min in the absence or presence of 100 nM JWH-133 or WIN-55,212-2. B, C6-cell gliomas. Mice were treated as in Fig. 1 ⇓ for 8 days, tumors were dissected, and immunomicroscopy analysis was performed. Abbreviations as in Fig. 1 ⇓ . Fig. 2b ⇓ . Regression of human astrocytomas in vivo upon selective CB2 receptor activation. A, example of CB2 receptor expression in a grade IV human astrocytoma (HA) as assessed by immunomicroscopy analysis of a tumor section and of cells derived from the tumor. Western blot analysis of the tumor was also performed using C6 glioma cells as a control. In B, cells from the tumor shown in A were injected s.c. in mice. When tumors had reached the desired size (day 0), animals were treated with either vehicle (?) or JWH-133 (•) for 25 days (n = 6 for each experimental group). C, examples of s.c. gliomas in the flank of mice (top panel) and after dissection (bottom panel) after treatment with vehicle or JWH-133 (JWH) for 25 days.
Regression of Human Astrocytomas in Vivo by Selective CB2 Receptor Activation.
The presence of CB1 and CB2 receptors was analyzed by immunohistochemistry in biopsies from human astrocytomas of low (grades I–II) and high (grades III–IV) malignancy. High malignancy tumors were diagnosed in all cases as pure astrocytomas, whereas some of the low malignancy tumors were mixed oligoastrocytomas. However, no differences were observed in the distribution of cannabinoid receptors between pure and mixed tumors (data not shown). A full 70% (26 of 37) of the total tumors analyzed expressed measurable levels of cannabinoid receptors (i.e., CB1 and/or CB2). Among them, a similar percentage of low malignancy (4 of 12) and high malignancy tumors (6 of 25) expressed CB1 and CB2 receptors simultaneously. Of interest, the extent of CB2 receptor expression was related with tumor malignancy. Thus, as shown in Table 1 ⇓ , the proportion of grade IV astrocytomas showing high/very high CB2 immunoreactivity doubled that of grades I–III tumors. Moreover, CB2 receptor expression was markedly enhanced in grade IV astrocytomas compared with CB1 receptor expression.
Given the inhibition of C6-cell glioma growth by selective CB2 receptor activation (Fig. 1) ⇓ , we evaluated the effect of JWH-133 treatment on the growth of highly malignant (grade IV) human astrocytoma cells in vivo. Immunofluorescence microscopy and Western blot analyses evidenced the expression of the CB2 receptor in the inoculated cells (Fig. 2b ⇓ , A). This particular tumor also expressed the CB1 receptor (data not shown). Of interest, JWH-133 administration completely blocked the proliferation of the human astrocytoma (Fig. 2b ⇓ , B). Examples of tumor-bearing mice and of dissected tumors after cannabinoid treatment for 25 days are shown in Fig. 2b ⇓ , C.
Selective CB2 Receptor Activation Signals Apoptosis of C6 Glioma Cells via Ceramide Synthesis de Novo.
We have shown previously that THC-induced apoptosis of C6 glioma cells relies on the sustained generation of the proapoptotic lipid ceramide (20 , 29) . To obtain further evidence for the specificity of the JWH-133 antitumoral action, experiments were carried out with L-cycloserine, a selective competitive inhibitor of SPT, the enzyme which catalyzes the pace-setting step of ceramide synthesis de novo (30) . As shown in Fig. 3, A and B ⇓ , exposure to JWH-133 induced apoptosis of C6 glioma cells, and this effect was prevented by L-cycloserine. Moreover, L-cycloserine was able to suppress JWH-133-evoked SPT induction (Fig. 3C) ⇓ . We also tested the effect of L-cycloserine on JWH-133-induced ERK activation, as ERK seems to be the downstream target of ceramide in THC-evoked apoptosis of C6 glioma cells (20 , 29) . Thus, blockade of ceramide synthesis de novo with L-cycloserine abrogated JWH-133-induced ERK activation (Fig. 3D) ⇓ , indicating that JWH-133, like THC, signals apoptosis via ceramide synthesis de novo and ERK activation.
Fig. 3. http://
Selective CB2 receptor activation induces apoptosis of C6 glioma cells via ceramide synthesis de novo. C6 glioma cells were cultured in the absence or presence of 100 nM JWH-133 (JWH) and/or 0.5 mM L-cycloserine (CS) for 5 days. A, TUNEL staining and phase-contrast micrographs. Images of a representative experiment are shown. Similar results were obtained in two other experiments. B, cell viability (n = 6). C, SPT activity (n = 4). D, ERK activity (n = 6). ∗, significantly different (P < 0.01) from incubations with no additions.
DISCUSSION
Recent studies have dealt with the antiproliferative effect of cannabinoids on different transformed cells. However, this property of cannabis compounds was first reported 25 years ago by Munson et al. (31) , who showed that cannabinoids may inhibit the growth of Lewis lung adenocarcinoma, B-tropic Friend leukemia virus-induced splenomegaly, and L1210 leukemia cells in vivo. Although these observations were actually promising, further investigations were not performed on this area until a few years ago. Nowadays, there are data demonstrating that cannabinoids inhibit the growth of transformed neural, breast, and prostate cells in culture (32) and induce the regression of gliomas in vivo (Ref. 20 and the present study). The case of gliomas is of particular interest because they are one of the most malignant forms of cancer, resulting in the death of affected patients within months after diagnosis. Conventional therapies, including surgery, radiotherapy, chemotherapy, and immunotherapy, are usually ineffective or just palliative (33 , 34) . The most recent strategies for glioma treatment are focused on gene therapy, but no trial performed thus far has been significantly successful (33, 34, 35) . It is therefore essential to develop new therapeutic strategies for the management of gliomas and most likely to combine some of them to obtain significant clinical results. One of these alternative therapeutic approaches might be on the basis of the use of nonpsychoactive cannabinoid ligands, because these compounds induce apoptosis in vitro and inhibition of tumor growth without significant collateral effects in vivo. Because of the current inability to kill target glioma cells with sufficient efficacy to obtain significant tumor regression in the clinical practice, the combined use of drugs that inhibit tumor growth with cytotoxic approaches may be envisaged as the most feasible anticancer strategy in the near future. Of interest, cannabinoid apoptotic action relies on the generation of ceramide, a lipid second messenger that may have antitumoral properties either alone (36) or in combined therapies (37) .
We are nevertheless aware that under certain circumstances, cannabinoids may be immunosuppressive compounds by acting on immune organs and cells via CB2 receptors, and this would be expected to inhibit host antitumor immunity. As a matter of fact, Zhu et al. (38) have recently reported that i.p. THC injection to immune-competent mice for 4–6 weeks leads to an accelerated growth of tumor implants in two different murine lung cancer models. This effect, although not evidenced in the former report by Munson et al. (31) , was shown to rely on the CB2-dependent inhibition of the capacity of antigen-presenting cells and T cells to generate alloreactivity (38) . It is therefore possible that cannabinoids exert a dual effect on tumor growth, i.e., a direct antiproliferative effect (Ref. 20 and the present study) and an indirect growth-enhancing effect via inhibition of immunogenicity (38) . Factors such as the route of drug administration (local versus systemic), the timing of drug delivery (short-term versus long-term treatment), and the intrinsic capacity of a particular tumor cell to respond to cannabinoids (e.g., presence versus absence of cannabinoid receptors) might determine the balance between tumor progression and regression. In any event, the present study, together with our previous observations (20) , shows that the antitumoral action of cannabinoids on gliomas may be exerted either via the CB1 receptor or via the CB2 receptor. The attractive possibility of finding cannabinoid-based therapeutic strategies for neural diseases devoid of nondesired CB1-mediated psychotropic side effects is also opened by the possible implication of the CB2 receptor in the control of pain initiation (12) and multiple sclerosis-linked spasticity (16) . Moreover, our data support the notion that the CB2 receptor might serve as a diagnostic marker of glial cell proliferation/malignancy, in line with what Valk et al. (39 , 40) have reported for myeloid cell growth and transformation during leukemogenesis.

Acknowledgments
We thank J. L. Gil, M. E. Fernández de Molina, C. Bailón, C. Hernández, and J. Palacín for their expert technical assistance. This article is dedicated to Dr. Math J. H. Geelen, outstanding on personal and scientific grounds, on his retirement as Professor of Veterinary Biochemistry.